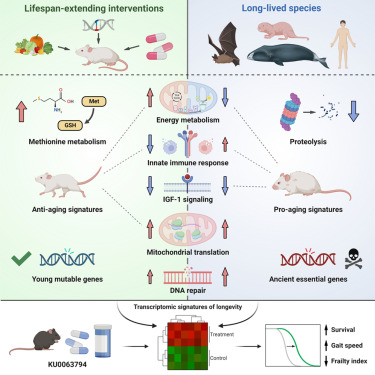
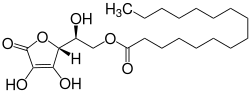
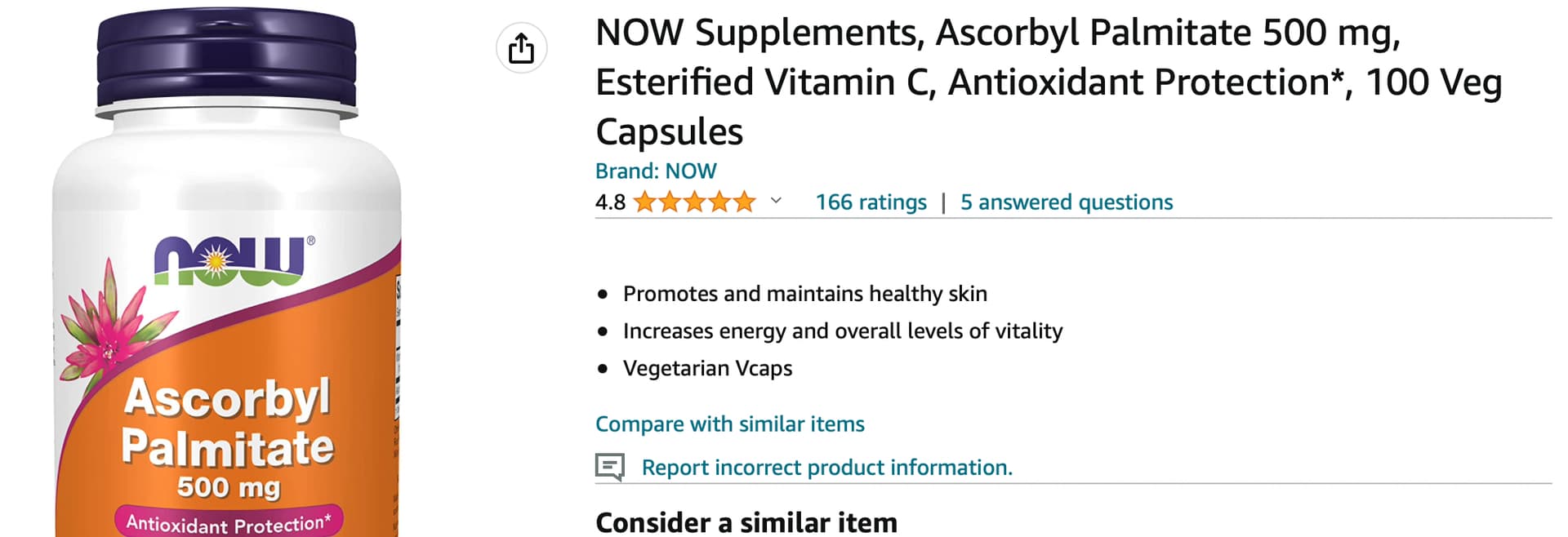
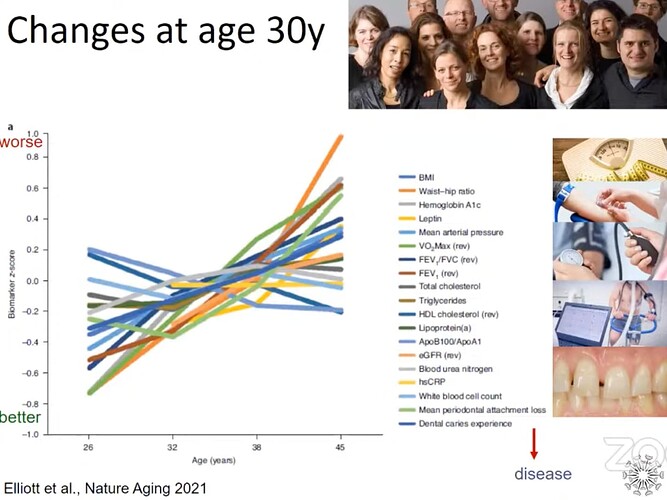
I’ve been taking it for a few years now… research looks positive, no obvious downsides:
See on Amazon:
https://www.amazon.com/gp/product/B001F0R66A/ref=ppx_yo_dt_b_search_asin_title?ie=UTF8&psc=1
Also - some interesting information on the other compounds mentioned:
AZD8055
The serine/threonine kinase mammalian target of rapamycin (mTOR) is crucial for cell growth and proliferation, and is constitutively activated in primary acute myeloid leukemia (AML) cells, therefore representing a major target for drug development in this disease. We show here that the specific mTOR kinase inhibitor AZD8055 blocked mTORC1 and mTORC2 signaling in AML. Particularly, AZD8055 fully inhibited multisite eIF4E-binding protein 1 phosphorylation, subsequently blocking protein translation, which was in contrast to the effects of rapamycin. In addition, the mTORC1-dependent PI3K/Akt feedback activation was fully abrogated in AZD8055-treated AML cells. Significantly, AZD8055 decreased AML blast cell proliferation and cell cycle progression, reduced the clonogenic growth of leukemic progenitors and induced caspase-dependent apoptosis in leukemic cells but not in normal immature CD34+ cells. Interestingly, AZD8055 strongly induced autophagy, which may be either protective or cell death inducing, depending on concentration. Finally, AZD8055 markedly increased the survival of AML transplanted mice through a significant reduction of tumor growth, without apparent toxicity. Our current results strongly suggest that AZD8055 should be tested in AML patients in clinical trials.
More here:
https://www.nature.com/articles/leu2011339
Preclinical pharmacology
AZD8055 is a potent, selective inhibitor of mTOR kinase, targeting both mTORC1 (rapamycin-sensitive) and mTORC2 (rapamycin insensitive) complexes. AZD8055 is specific against mTOR (IC50 value of 0.8 ± 0.2 nM using an immunoprecipitate of full length mTOR from HeLa cells in an ELISA-based kinase assay). AZD8055 was inactive in a counter screen against 260 kinases.
AZD8055 inhibited downstream targets of both mTORC1 (phosphorylation of S6 at serine 235/236) and mTORC2 (phosphorylation of AKT at serine 473) in several in vitro models in a dose- and time-dependent manner. Oral treatment of mice bearing U87-MG human glioma xenografts twice daily with 2.5, 5, and 10 mg/kg/day AZD8055 for 10-days resulted in a dose-dependent tumour growth inhibition of 33%, 48% and 77%, respectively. All doses were well tolerated. The growth inhibitory effect of once daily administration with 10 and 20 mg/kg of AZD8055 for 10-days was 57 and 85%, respectively. This tumour growth inhibitory effect was observed in a number of xenograft models tested in vivo. Furthermore, the antitumour activity in vivo was in good agreement with growth inhibition observed in cell lines in vitro.
More here:
https://openinnovation.astrazeneca.com/preclinical-research/preclinical-molecules/azd8055.html
GDC0941 / Pictilisib
Purpose
This first-in-human dose-escalation trial evaluated the safety, tolerability, maximal tolerated dose (MTD), dose limiting toxicities (DLTs), pharmacokinetics, pharmacodynamics and preliminary clinical activity of pictilisib (GDC-0941), an oral, potent and selective inhibitor of the Class I phosphatidylinositol-3-kinases (PI3K)
Results
Pictilisib was well-tolerated. The most common toxicities were grade 1-2 nausea, rash and fatigue while the DLT was grade 3 maculopapular rash (450mg, 2 of 3 patients; 330mg, 1 of 7 patients). The pharmacokinetic profile was dose-proportional and supported once-daily dosing. Levels of phosphorylated serine-473 AKT were suppressed >90% in platelet rich plasma at 3 hours post-dose at the MTD and in tumor at pictilisib doses associated with AUC >20uM.hr. Significant increase in plasma insulin and glucose levels, and >25% decrease in 18F-FDG uptake by PET in 7 of 32 evaluable patients confirmed target modulation. A patient with V600E BRAF mutant melanoma and another with platinum-refractory epithelial ovarian cancer exhibiting PTEN loss and PIK3CA amplification demonstrated partial response by RECIST and GCIG-CA125 criteria, respectively.
Conclusion
Pictilisib was safely administered with a dose-proportional pharmacokinetic profile, on-target pharmacodynamic activity at dose levels ≥100mg and signs of antitumor activity. The recommended Phase II dose was continuous dosing at 330mg once-daily.
A Phase 2 trial with another drug/combination:
Conclusion
Adding pictilisib to anastrozole significantly increases suppression of tumor cell proliferation in luminal B primary breast cancer.
Phase II Randomized Preoperative Window-of-Opportunity Study of the PI3K Inhibitor Pictilisib Plus Anastrozole Compared With Anastrozole Alone in Patients With Estrogen Receptor–Positive Breast Cancer
https://ascopubs.org/doi/10.1200/jco.2015.63.9179
Availabilty and Pricing: