Fantastic reporting … thanks for sharing!
A few papers I could dig up on Thioredoxin reductase and GPLD1
Mitochondrial thioredoxin reductase 2 is elevated in long-lived primate as well as rodent species and extends fly mean lifespan
In a survey of enzymes related to protein oxidation and cellular redox state, we found activity of the redox enzyme thioredoxin reductase (TXNRD) to be elevated in cells from long-lived species of rodents, primates, and birds. Elevated TXNRD activity in long-lived species reflected increases in the mitochondrial form, TXNRD2, rather than the cytosolic forms TXNRD1 and TXNRD3. Analysis of published RNA-Seq data showed elevated TXNRD2 mRNA in multiple organs of longer-lived primates, suggesting that the phenomenon is not limited to skin-derived fibroblasts. Elevation of TXNRD2 activity and protein levels was also noted in liver of three different long-lived mutant mice, and in normal male mice treated with a drug that extends lifespan in males. Overexpression of mitochondrial TXNRD2 in Drosophila melanogaster extended median (but not maximum) lifespan in female flies with a small lifespan extension in males; in contrast, overexpression of the cytosolic form, TXNRD1, did not produce a lifespan extension.
Thioredoxin and thioredoxin reductase are evolutionarily conserved antioxidant enzymes that protect organisms from oxidative stress. These proteins also play roles in redox signaling and can act as a redox-independent cellular chaperone. In most organisms, there is a cytoplasmic and mitochondrial thioredoxin system. A number of studies have examined the role of thioredoxin and thioredoxin reductase in determining longevity. Disruption of either thioredoxin or thioredoxin reductase is sufficient to shorten lifespan in model organisms including yeast, worms, flies and mice, thereby indicating conservation across species. Similarly, increasing the expression of thioredoxin or thioredoxin reductase can extend longevity in multiple model organisms. In humans, there is an association between a specific genetic variant of thioredoxin reductase and lifespan. Overall, the cytoplasmic and mitochondrial thioredoxin systems are both important for longevity.
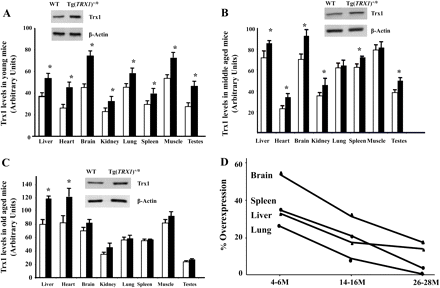
Thioredoxin 1 Overexpression Extends Mainly the Earlier Part of Life Span in Mice
We examined the effects of increased levels of thioredoxin 1 (Trx1) on resistance to oxidative stress and aging in transgenic mice overexpressing Trx1 [Tg(TRX1 )+/0]. The Tg(TRX1 )+/0 mice showed significantly higher Trx1 protein levels in all the tissues examined compared with the wild-type littermates. Oxidative damage to proteins and levels of lipid peroxidation were significantly lower in the livers of Tg(TRX1 )+/0 mice compared with wild-type littermates. The survival study demonstrated that male Tg(TRX1 )+/0 mice significantly extended the earlier part of life span compared with wild-type littermates, but no significant life extension was observed in females. Neither male nor female Tg(TRX1 )+/0 mice showed changes in maximum life span. Our findings suggested that the increased levels of Trx1 in the Tg(TRX1 )+/0 mice were correlated to increased resistance to oxidative stress, which could be beneficial in the earlier part of life span but not the maximum life span in the C57BL/6 mice.
And GPLD1
Cap-independent translation of GPLD1 enhances markers of brain health in long-lived mutant and drug-treated mice
Glycosylphosphatidylinositol-specific phospholipase D1 (GPLD1) hydrolyzes inositol phosphate linkages in proteins anchored to the cell membrane. Mice overexpressing GPLD1 show enhanced neurogenesis and cognition. Snell dwarf (DW) and growth hormone receptor knockout (GKO) mice show delays in age-dependent cognitive decline. We hypothesized that augmented GPLD1 might contribute to retained cognitive function in these mice. We report that DW and GKO show higher GPLD1 levels in the liver and plasma. These mice also have elevated levels of hippocampal brain-derived neurotrophic factor (BDNF) and of doublecortin (DCX), suggesting a mechanism for maintenance of cognitive function at older ages. GPLD1 was not increased in the hippocampus of DW or GKO mice, suggesting that plasma GPLD1 increases elevated these brain proteins. Alteration of the liver and plasma GPLD1 was unaltered in mice with liver-specific GHR deletion, suggesting that the GH effect was not intrinsic to the liver. GPLD1 was also induced by caloric restriction and by each of four drugs that extend lifespan. The proteome of DW and GKO mice is molded by selective translation of mRNAs, involving cap-independent translation (CIT) of mRNAs marked by N6 methyladenosine. Because GPLD1 protein increases were independent of the mRNA level, we tested the idea that GPLD1 might be regulated by CIT. 4EGI-1, which enhances CIT, increased GPLD1 protein without changes in GPLD1 mRNA in cultured fibroblasts and mice. Furthermore, transgenic overexpression of YTHDF1, which promotes CIT by reading m6A signals, also led to increased GPLD1 protein, showing that elevation of GPLD1 reflects selective mRNA translation.
Lifespan extension in female mice by early, transient exposure to adult female olfactory cues
Several previous lines of research have suggested, indirectly, that mouse lifespan is particularly susceptible to endocrine or nutritional signals in the first few weeks of life, as tested by manipulations of litter size, growth hormone levels, or mutations with effects specifically on early-life growth rate. The pace of early development in mice can also be influenced by exposure of nursing and weanling mice to olfactory cues. In particular, odors of same-sex adult mice can in some circumstances delay maturation. We hypothesized that olfactory information might also have a sex-specific effect on lifespan, and we show here that the lifespan of female mice can be increased significantly by odors from adult females administered transiently, that is from 3 days until 60 days of age. Female lifespan was not modified by male odors, nor was male lifespan susceptible to odors from adults of either sex. Conditional deletion of the G protein Gαo in the olfactory system, which leads to impaired accessory olfactory system function and blunted reproductive priming responses to male odors in females, did not modify the effect of female odors on female lifespan. Our data provide support for the idea that very young mice are susceptible to influences that can have long-lasting effects on health maintenance in later life, and provide a potential example of lifespan extension by olfactory cues in mice.
Blood-to-brain communication in aging and rejuvenation
Aging induces molecular, cellular and functional changes in the adult brain that drive cognitive decline and increase vulnerability to dementia-related neurodegenerative diseases. Leveraging systemic and lifestyle interventions, such as heterochronic parabiosis, administration of ‘young blood’, exercise and caloric restriction, has challenged prevalent views of brain aging as a rigid process and has demonstrated that aging-associated cognitive and cellular impairments can be restored to more youthful levels. Technological advances in proteomic and transcriptomic analyses have further facilitated investigations into the functional impact of intertissue communication on brain aging and have led to the identification of a growing number of pro-aging and pro-youthful factors in blood. In this review, we discuss blood-to-brain communication from a systems physiology perspective with an emphasis on blood-derived signals as potent drivers of both age-related brain dysfunction and brain rejuvenation.
https://www.nature.com/articles/s41593-022-01238-8
It Seems that GPLD1 is available as a research molecule: