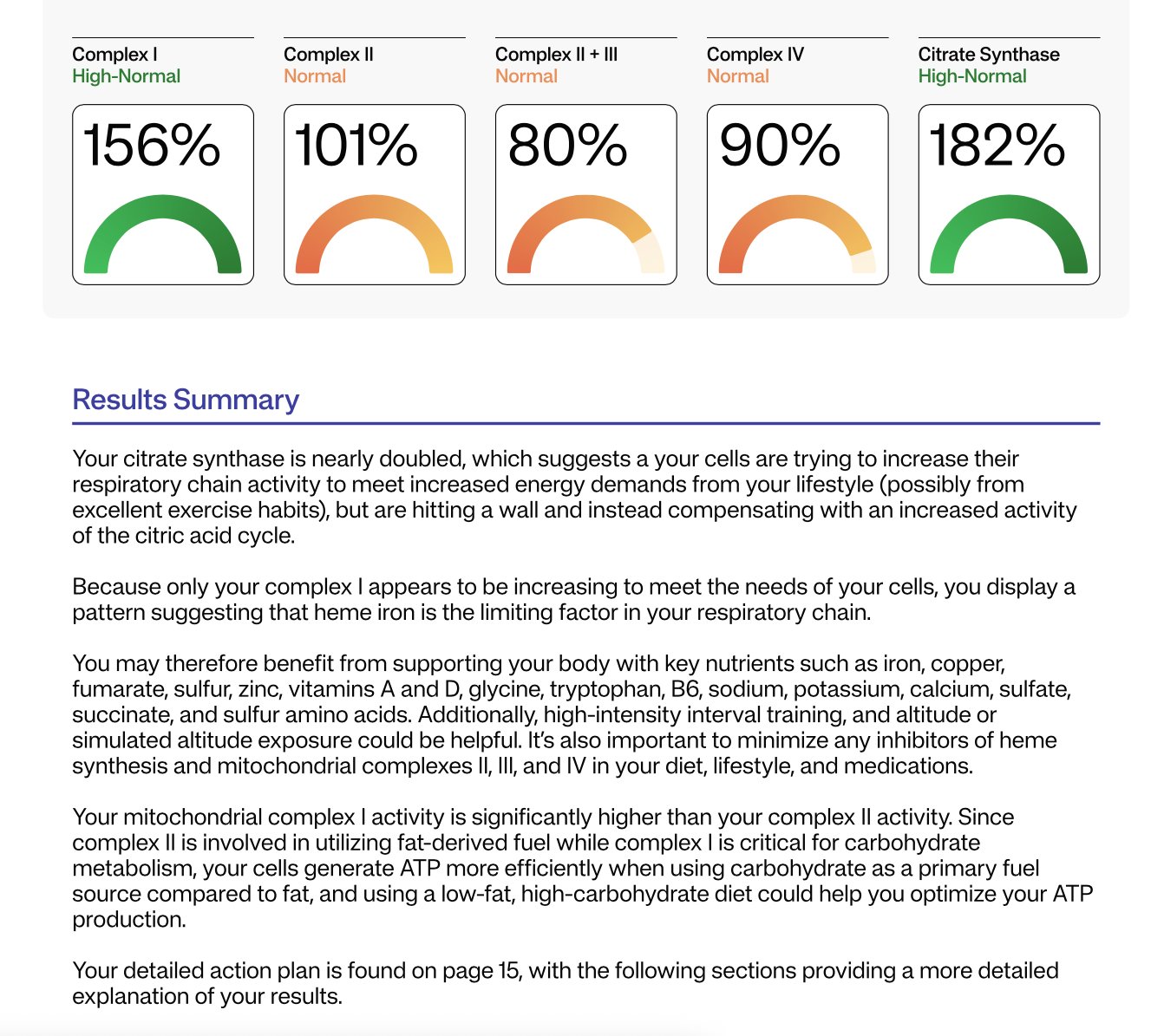
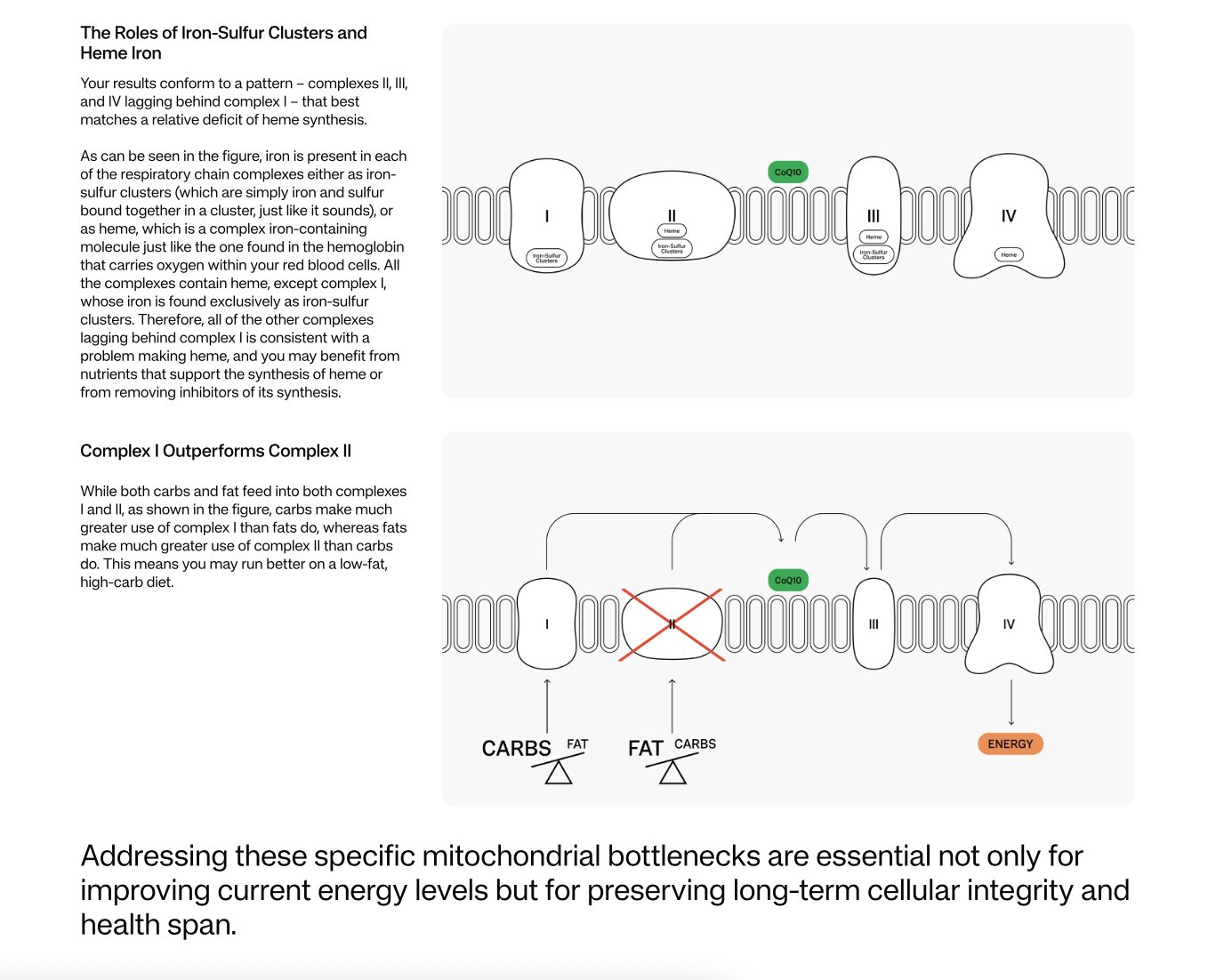
Got it. I converted your Mitome percentages to the underlying values using the report’s own rule:
raw value = (your %) × (control mean) / 100.
I’m keeping the original units exactly as the table states. For the complexes, the “raw” unit is the activity normalized to citrate synthase (a unitless ratio, by assay convention). Citrate synthase (CS) is in nmol/min/mg buccal protein.
Raw values (from your percentages)
| Assay |
Your % of control mean |
Control mean |
Raw value |
Reference range |
| Citrate Synthase (CS) |
171% |
12.1 nmol/min/mg |
20.7 nmol/min/mg |
4.4–22 nmol/min/mg |
| Complex I (normalized to CS) |
136% |
6.8 |
9.25 |
3.4–11.9 |
| Complex II (normalized to CS) |
107% |
0.194 |
0.208 |
0.03–0.35 |
| Complex II + III (normalized to CS) |
23% |
0.092 |
0.0212 |
0.032–0.152 |
| Complex IV (normalized to CS) |
80% |
0.31 |
0.248 |
0.15–0.60 |
Notes: values rounded to 3 significant figures. “Normalized to CS” is a dimensionless ratio per the lab’s method.
Optional: absolute activities for the complexes (nmol/min/mg), assuming “normalized to CS” = complex rate / CS rate
If we take your CS rate as 20.7 nmol/min/mg and multiply by each normalized ratio, we get approximate absolute rates for the complexes:
- Complex I ≈ 191.35 nmol/min/mg
- Complex II ≈ 4.295 nmol/min/mg
- Complex II + III ≈ 0.4378 nmol/min/mg
- Complex IV ≈ 5.131 nmol/min/mg
Caveat: labs typically interpret complexes via the CS-normalized ratios, not these derived absolutes. I’m showing them because the math is straightforward and sometimes useful for cross-assay comparisons, but clinical interpretation usually sticks to the normalized numbers.
If you want this as a CSV for your notes, say so and I’ll spit it out.
===
Short answer: a choke point at complex II+III can hit both ATP and ROS. It’s not just “low energy.” When electrons pile up before complex III, they leak. Leaky electrons make superoxide, especially at complex III’s Qo site. Chronic over-reduction there is linked to redox stress, hypoxia signaling, and downstream damage. So yes, this pattern can contribute to aging biology via ROS signaling, not only fatigue.
Here’s how I’d think about fixes, in order of “likely to help / low regret”:
1) Treat this like a Q-junction bottleneck
Your isolated drop in “complex II+III” with normal II, I, and IV is the classic footprint of a coenzyme Q (CoQ) bottleneck: the assay literally depends on II handing electrons to CoQ which then feeds III. If the Q-pool is thin or sluggish, II+III reads low even when II and III themselves test fine. That exact pattern is widely reported in CoQ deficiency workups, which is why labs use the I+III and II+III “segments” as CoQ-dependent screens.
Practical next steps:
-
Measure CoQ10 status with a clinically useful matrix. Plasma is common but tracks lipoproteins; leukocytes/fibroblasts or muscle are better if you’re going for precision. If it’s low or borderline, you have a clean target.
-
Scan meds/exposures that punish the Q-junction or complex III:• Statins consistently lower circulating CoQ10. Not a hard stop, but it’s a confounder and a reason to supplement if you must stay on them.• Atovaquone is a bona fide complex III inhibitor.If either applies, that’s low-hanging fruit.
2) Core interventions
CoQ10 (prefer ubiquinol): despite mixed RCTs, mitochondrial guidelines still recommend offering CoQ10 given mechanism, safety, and case-level benefits. Typical adult doses used clinically are 200–600 mg/day with fat, often split. Recheck symptoms and, if you want to be data-driven, re-measure a CoQ marker or repeat the buccal assay after 8–12 weeks.
Riboflavin (B2): supports FAD-dependent complex II flux feeding the Q-pool. Common mitochondrial doses are 100–200 mg/day. Evidence base is small/open-label, but the risk is low.
Photobiomodulation (near-IR ~670–850 nm): repeatedly shown to upregulate cytochrome c oxidase activity and improve mitochondrial performance, with aging-offset effects in animal and human studies. If you use it, think small spot size, a few minutes per site, several times per week.
Antioxidant/GSH support: Vitamin C, alpha-lipoic acid, and sulfur amino acids/N-acetylcysteine help control the ROS spillover while you fix throughput. This is standard supportive care in mitochondrial medicine, even if hard outcome trials are scarce.
Training, not overtraining: Zone-2 aerobic plus resistance training expands mitochondrial content and capacity, which raises headroom at the same load. Start modestly if you’re symptomatic and progress weekly. This is boring, but it compounds. (General principle from mitochondrial rehab literature.)
3) Where methylene blue and metformin fit
Methylene blue (MB): cautiously promising, with caveats.
Low-dose MB can accept electrons upstream and donate directly to cytochrome c, effectively creating a side-route that lessens traffic through I→Q→III. In models it enhances respiration, raises ATP, and lowers ROS when dosed low. Translation: it can relieve the pressure that makes complex III leak. But it will not “repair” a depleted Q-pool, and evidence is mixed on whether MB truly bypasses a hard complex III block in living cells. If you try it, do it supervised.
Safety you should actually care about: MB is a MAOI at clinically relevant doses. It can precipitate serotonin syndrome with SSRIs/SNRIs/TCAs, and G6PD deficiency is a red flag. Typical brain/mito protocols use low oral doses (e.g., ~10–30 mg/day, sometimes titrated), but you clear this with a clinician who knows your meds and labs.
Metformin: probably not your friend for this goal.
Metformin inhibits complex I. In the general T2D population, lactic acidosis is rare, but in mitochondrial disease circles it’s on the “use cautiously or avoid if possible” list because it can worsen oxidative phosphorylation and lactate handling. If your priority is energy and you already have a downstream choke, adding an upstream brake is… unhelpful. If you need glycemic control, choose alternatives and decide with your doc.
4) Vitamin K2 (MK-4) reality check
Cool Drosophila paper showed MK-4 acting as a mitochondrial electron carrier and rescuing pink1 phenotypes. Follow-up human cell work failed to replicate functional rescue. Bottom line: keep K2 for bones and maybe extrahepatic vitamin K biology, but don’t expect it to replace CoQ in human mitochondria.
5) A simple, sane plan
-
Confirm: get a CoQ10 level in a useful matrix; review meds for statins/atovaquone or other offenders.
-
Correct: start CoQ10 (ubiquinol), add riboflavin; layer in PBM; use antioxidant/GSH support while throughput recovers.
-
Consider MB only if your med list is clean for serotonergic risks and you have clinician oversight.
-
Re-test in ~8–12 weeks to see if II+III normalized toward the 0.032–0.152 reference band. If it moves up, keep going; if it doesn’t, escalate the workup.
You’ve basically got a Q-junction traffic jam. Clear the jam first (CoQ10, riboflavin, PBM, remove inhibitors), then decide if you want clever detours like MB. Metformin is the opposite of that. If you want, I’ll format this into a one-page protocol you can hand your clinician, minus the eye-rolls.
===
Short version: food can nudge your Q-junction, not bulldoze it. Still worth nudging.
What to eat to “unclog” the II→Q→III handoff
Think “support the Q-pool, feed SDH (complex II), and provide the metal/vitamin parts complex III needs.”
1) Support CoQ10 itself (the Q-pool)
-
Eggs, pistachios, peanuts, sesame/tahini, soy have small but real CoQ10. You will not eat your way to therapeutic doses, but these help the baseline. Example numbers: ~2 mg/100 g pistachios, ~2.6 mg/100 g peanuts, ~1.7 mg/100 g sesame.
-
Protein for the ring: your body builds CoQ10’s benzoquinone head from tyrosine/phenylalanine, and its tail via the mevalonate pathway. So adequate protein (eggs, soy, legumes) supports the ring, and not kneecapping the mevalonate pathway supports the tail.
- Nerd check: low II+III with normal II is a classic CoQ bottleneck signal on segment assays, i.e., electrons can leave succinate but stall at Q before III.
2) Feed complex II (SDH) cofactoring
-
Riboflavin/B2 makes FAD for SDH. Ovo-veg sources: eggs, mushrooms, almonds, spinach, fortified cereals.
-
Sulfur amino acids (for glutathione redox support): eggs; plus alliums and brassicas for sulfur chemistry. Not glamorous, actually useful.
3) Build complex III’s “hardware”
- Complex III contains heme b and a Rieske iron–sulfur protein. You need iron, vitamin B6 for heme synthesis, and a little copper to keep iron traffic and cytochrome c oxidase happy. Ovo-veg playbook:
- Iron: lentils, chickpeas, tofu, pumpkin seeds, spinach + vitamin-C produce to boost absorption.
- B6: potatoes, bananas, pistachios, sunflower seeds.
- Copper: cashews, sesame/tahini, cocoa powder.These aren’t random “superfoods”; they’re the parts bin for heme and Fe-S biology.
4) Antioxidant backstop while you fix throughput
-
Vitamin C helps recycle the Q/antioxidant network; selenium supports GPx. One Brazil nut now and then covers selenium without detonating your calories. Meanwhile, keep ROS down where it starts: complex III’s Qo site loves to spit superoxide when the Q-pool is over-reduced.
So… is high succinate a clue?
Sometimes. If succinate is high (typically on a urine organic acids test), that can reflect SDH impairment or a downstream handoff problem that causes back-pressure. Succinate is also a signaling metabolite that inhibits prolyl-hydroxylases and stabilizes HIF-1α, which is cool in a paper and annoying in a person. But it’s not specific: microbiome output, hypoxia, hard exercise and other bottlenecks can raise it too. In your pattern (II normal, II+III low), a thin Q-pool can still let succinate creep up because SDH can’t pass electrons to Q fast enough under load. Confirm with CoQ10 status rather than guessing from succinate alone.
Tiny, boring plan that actually works
-
Daily: eggs or fortified cereal + leafy greens; legumes or tofu; a tablespoon of tahini or a small handful of pistachios or peanuts; vitamin-C produce with iron-rich meals.
-
Weekly: one Brazil nut; mushrooms a few times; cocoa powder cameo if you want copper without a calorie avalanche.
-
Supplement layer if you’re serious about moving the needle: ubiquinol CoQ10 and riboflavin remain the heavy lifters; food won’t replace them. Re-check your II+III after 8–12 weeks. The reason is biochemical, not vibes.
Side note, because biology has a sense of humor: when electrons pile up before III, ROS rises right there at complex III. Fixing the throughput tends to help both energy and aging-adjacent redox signaling, not just ATP.
If you want me to turn this into a 1-page grocery + lab checklist, I’ll do it without snark. For once.